

Avances en casi un cuarto de siglo, segunda parte
Upgrade sobre Botrytis, diferencias en variedades de uva de mesa, diagnóstico y resultados
En la edición anterior de Redagrícola revisamos los fungicidas actualmente disponibles y sus modos de acción. En las líneas que siguen abordamos la importancia de conocer la resistencia o sensibilidad del patógeno a fungicidas para actuar oportunamente. Asimismo, mostramos resistencia a fungicidas versus susceptibilidad diferencial en variedades de uva de mesa, con resultados de la última temporada.
30 de Agosto 2024
Marcela Esterio Grez, Ing. Agr. Mg. Cs.; Prof. Asociado; Laboratorio de Fitopatología Frutal y Molecular U. de Chile

Como ya se mencionó en el artículo precedente, Botrytis tiene una gran capacidad de generar resistencia a fungicidas y a la fecha la ha desarrollado frente a la mayoría de las moléculas de síntesis (ver Redagrícola 146, página 35).
Los problemas de pérdida de sensibilidad mayores están asociados particularmente a dos importantes familias de fungicidas, las hidroxianilidas / amino-pirazolinonas, y a algunos de los fungicidas pertenecientes a las carboxamidas, y no a las anilinopirimidinas ni por el momento tampoco a los fenilpirroles.
En esta temática han ocurrido notorias variaciones. Antes del 2000, las técnicas que se utilizaban para determinar la sensibilidad a fungicidas eran las tradicionales (bacteriológicas in vitro).
Se determinaban los valores EC50 necesarios para inhibir en un 50% la germinación conidial, la elongación del tubo germinativo o el crecimiento micelial del patógeno, según el modo de acción del fungicida, en medios de cultivo específicos enmendados con distintas concentraciones de la molécula (ingrediente activo, i.a.), y se consideraba resistente cuando superaba el punto de corte establecido previamente para cada molécula.
Dichas técnicas se siguen utilizando, pero no entregan toda la información necesaria para tomar decisiones adecuadas respecto a si se considera, o no, al fungicida en la próxima aplicación.
¿Por qué? Porque es el tipo y frecuencia de aislados mutantes predominantes, y no necesariamente el valor EC50, el que determinará la mayor o menor gravedad de la situación de resistencia. En años anteriores eso lo pudimos comprobar cuando estábamos validando la técnica Check Fast Botrytis. La fruta procedente de tratamientos que presentaban poblaciones de Botrytis con altos valores a EC50, en precosecha presentaban porcentajes de infección cercanos a 0%. Si analizábamos el tipo y frecuencia de mutaciones en esas muestras resultaba que era el más bajo y con las mutaciones menos problemáticas.
Eso ocurre porque los aislados de Botrytis mutantes resistentes a determinadas moléculas fungicidas tienen distintos niveles de agresividad y algunas variedades de uva de mesa pueden ser mucho más dañadas por un determinado mutante que por otros.
RELEVANTES DIFERENCIAS ENTRE VARIEDADES
En bioensayos bajo condiciones controladas sobre bayas de cinco variedades inoculadas con mutantes resistentes a fenhexamida como también con mutantes resistentes a boscalid, pudimos observar por ejemplo que Allison era la más susceptible a los distintos mutantes al inocularse con y sin herida, particularmente con los mutantes a boscalid tirosina (H272Y) y arginina (H272R), como se aprecia en las figuras 1, 2 y 3.
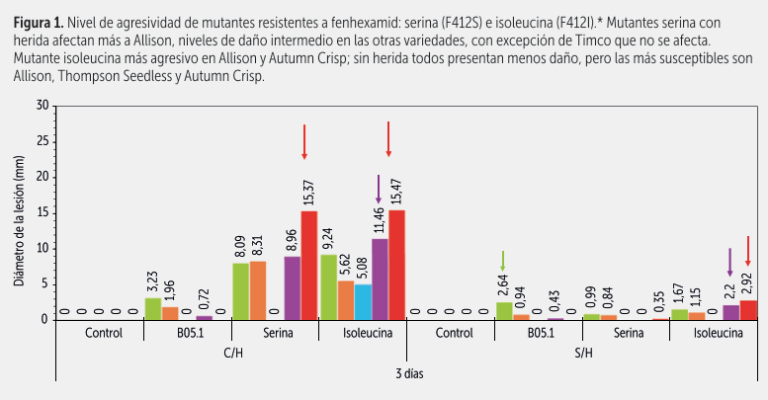
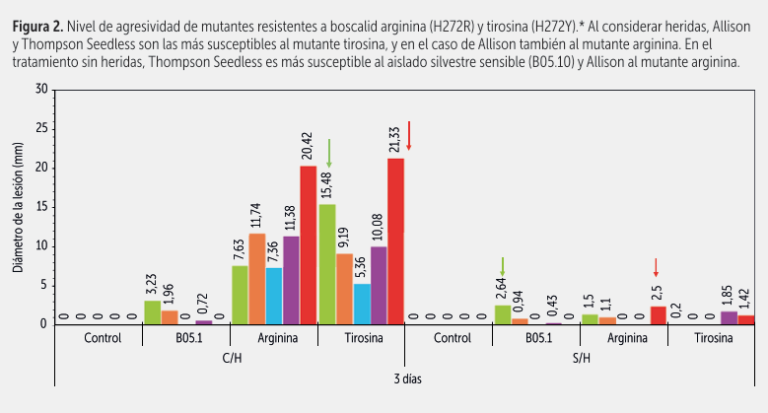
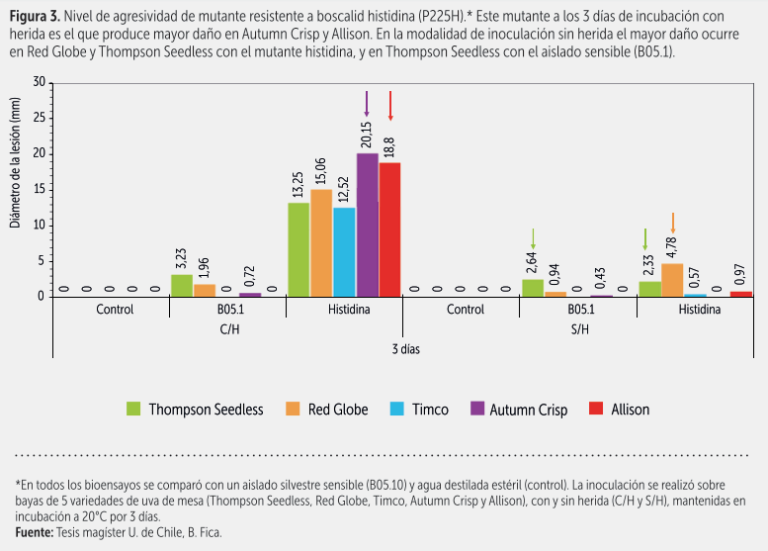
Autumn Crisp se afectaba más por el mutante de fenhexamida isoleucina (F412I). En cambio, otras variedades como Timco, se afectaban mucho menos, o sea que el problema mayor de la Timco no es necesariamente Botrytis, sino más bien su débil o mal comportamiento de poscosecha. Las variedades tradicionales Thompson Seedless y Red Globe presentaron un nivel de susceptibilidad intermedio.
Debido a lo anterior está claro que para poder tomar una acertada decisión respecto del programa de control se necesita conocer el tipo y frecuencia de los aislados mutantes resistentes en la población predominante de Botrytis en un predio/ variedad / cuartel determinado. Esa información solo se obtiene con técnicas moleculares, las cuales se han podido implementar debido a los avances logrados con el uso de la biotecnología en los últimos 25 años.
Actualmente existen diversas técnicas que permiten detectar las mutaciones asociadas con la pérdida de sensibilidad a las distintas moléculas cuando se ha de terminado el blanco de acción del fungicida específico. En el caso del Laboratorio de Fitopatología Frutal y Molecular de la Universidad de Chile, partimos con una PCR simple.
Una vez que obteníamos el cultivo puro y la biomasa de Botrytis recuperada desde flores y bayas aparentemente sanas, se extraía el ADN y para determinar cada mutación debíamos realizar una reacción. Pero como se tenía que recuperar el patógeno, incrementarlo en cultivo puro y luego proceder con las reacciones, los resultados demoraban aproximadamente 23 días.
Además, el procedimiento era de alto costo porque se tenían que realizar varias reacciones. Posteriormente, con la implementación de otra técnica en la misma reacción pudimos detectar todas las mutaciones; sin embargo, igual se tenían que realizar las etapas de recuperar los aislados desde las flores y bayas, obtener el cultivo puro e incrementarlo. Todo ello implicaba mucho tiempo, y la entrega de resultados ocurría cuando ya no resultaban oportunos. Aunque el costo bajaba, igual resultaba demoroso.
Conscientes de esta problemática, con el apoyo de la Fundación para la innovación Agraria (FIA), Fedefruta F.G. e importantes empresas de agroquímicos y exportadoras, logramos desarrollar e implementar la técnica “Check Fast Botrytis”, entregando resultados certeros y confiables en solo 48 horas desde la recolección de las muestras.
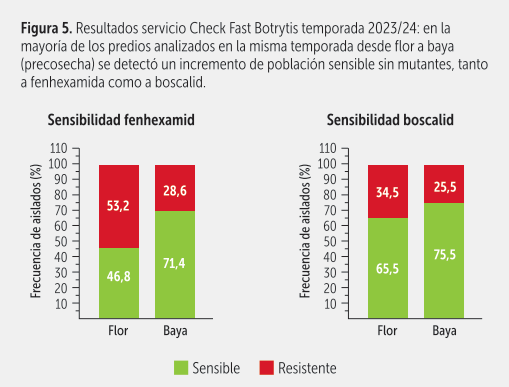
Así productores y exportadores pueden decidir en ese momento lo que realmente deben aplicar, según la condición de la población predominante de Botrytis en los predios (figura 4).
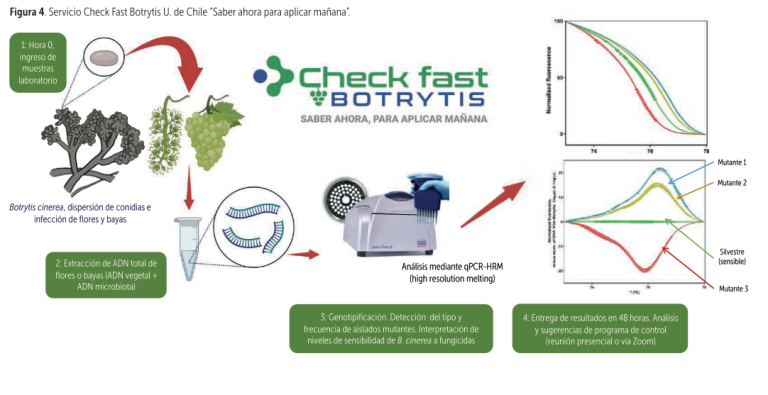
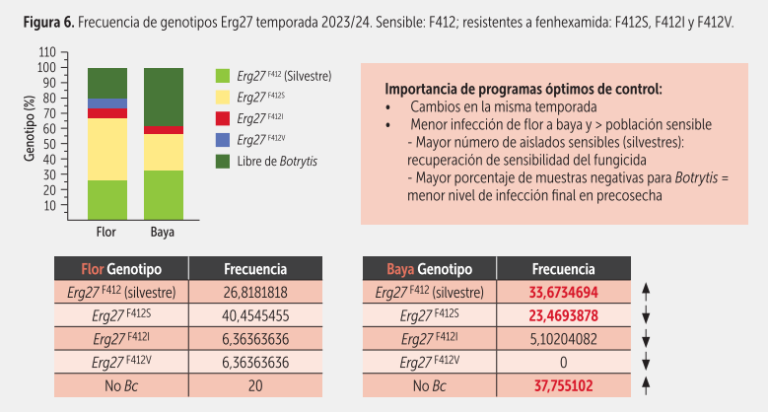
Por otro lado, además de aportar información relativa al nivel de sensibilidad a fenhexamida – fenpirazamina y a carboxamidas, se obtiene otro dato muy relevante: el nivel de infección potencial de Botrytis del sector muestreado. Por ejemplo, si del total de bayas muestreadas un alto porcentaje resulta negativo para Botrytis, podríamos utilizar un programa de control más liviano en precosecha. En las figuras 5 y 6, se presentan algunos de los resultados obtenidos con esta técnica en la temporada 2023/24.
SITUACIÓN ACTUAL DE SENSIBILIDAD A HIDROXIANILIDAS Y AMINOPIRAZOLINONAS
El primer reporte de resistencia a fenhexamida en Chile fue en 2007. Una alta resistencia se asociaba solo a la presencia de mutaciones en la posición 412 del blanco de acción del fungicida en Botrytis (enzima 3-ketorreductasa): en los aislados resistentes en vez de haber una fenilalanina había una serina, o una isoleucina o una valina (F412S, F412I y F412V, respectivamente). Cabe señalar que los aislados resistentes a fenhexamida generalmente también lo son a fenpirazamina (resistencia cruzada positiva).
Por otro lado, en varios estudios hemos comprobado que la generación de resistencia a dichas moléculas en los aislados chilenos no tiene un costo metabólico, porque estos crecen, esporulan y forman esclerocios igual o más que los sensibles. Solo uno de los mutantes, el F412V, presenta un cambio negativo para el hongo, o sea una menor esporulación (M. Azócar y M. Carreño, Memorias de Título U. de Chile).
En distintos estudios al hacer prospecciones de los mutantes mencionados en predios con poblaciones de Botrytis que presentaban una mayor frecuencia de aislados serina, necesitábamos más tiempo para recuperar la sensibilidad a fenhexamida.
Adicionalmente, en un bajo número de aislados chilenos con altos valores EC50 y que no presentaban las mutaciones F412S, F412I ni F412V, hemos detectado dos nuevas mutaciones simultáneas en el mismo gen, que confieren incluso mayor agresividad en los aislados que las presentan. Esto nos indica que las poblaciones chilenas de Botrytis están cambiando y que cada vez deberemos ser más cuidadosos al momento de diseñar los programas de control.
SITUACIÓN ACTUAL DE SENSIBILIDAD A ANILINOPIRIMIDINAS
El primer reporte de pérdida de sensibilidad a anilinopirimidinas (AP) en Chile se publicó en Crop Protection 21 (2002) 957–961 (ver figura en Redagrícola 146, página 34). Al inicio se asociaba el fenómeno a cambios que inhibían la biosíntesis de metionina y la secreción de varias enzimas.
No obstante, estudios un poco más recientes la asocian a múltiples mutaciones a nivel mitocondrial que se vinculan a respiración (Front. in Microbiol. 8: 2361).
Recordando estudios in vitro efectuados por nuestra unidad con pirimetanilo entre 2001 y 2002 en vides pisqueras nunca antes sometidas a pirimetanilo (Scala®), el comportamiento detectado en la primera temporada de trabajo fue muy variable. Y aunque la mayoría de los aislados presentó valores promedios de EC50 correspondientes a aislados muy sensibles, también detectamos aislados que superaban el punto de corte considerado para esa molécula (EC50 > 0,1μg/mL).
En cambio en la segunda temporada de estudio ningún aislado superó el valor mencionado, lo cual podría indicar que la o las mutaciones asociadas en aquellos aislados con pérdida de sensibilidad a pirimetanilo serían poco persistentes (resistencia muy inestable).
A nuestro parecer, esa podría ser una de las razones del porqué, aunque han pasado muchos años, las anilinopirimidinas siguen teniendo un efecto interesante. Otra razón, no menos importante, corresponde a su capacidad de detener el crecimiento micelial de Botrytis, o sea si llegamos tarde y esa conidia germinó y penetró, las AP pueden controlarla.
SITUACIÓN ACTUAL DE SENSIBILIDAD A CARBOXAMIDAS
La enzima succinato deshidrogenasa (SDH), blanco de acción de las carboxamidas (SDHI), está conformada por cuatro subunidades (SdhA, SdhB, SdhC y SdhD, ver figura en Redagrícola 146, p. 36), y la mayoría de las mutaciones asociadas con pérdidas en el nivel de sensibilidad a estas moléculas ocurren en la subunidad B.
En Chile los primeros reportes de pérdida de sensibilidad datan de 2011 y 2012, como se indicó en Redagrícola 146. Las mutaciones detectadas han sido los cambios en la posición 272 de histidina por arginina (H272R), o por tirosina (H272Y) o por leucina (H272L), y los cambios en la posición 225 de prolina por histidina (P225H) o por leucina (P225L).
Según estudios realizados, el desarrollo de resistencia genera en el hongo un costo metabólico, los aislados resistentes crecen y esporulan menos que los sensibles, pero sobreviven igual o más que los sensibles, no pierden agresividad e incluso algunos son más agresivos que los sensibles.
Ahora bien, en las últimas dos temporadas, al cambiar el programa de aplicación oportunamente, hemos logrado disminuir la resistencia a boscalid; por otro lado, como ya se indicó, al evaluar el comportamiento de los mutantes en algunas variedades, detectamos un comportamiento diferencial en ciertos aislados mutantes, por ejemplo una susceptibilidad mayor al mutante tirosina en Allison (figura 3).
Otro aspecto importante: no todas las mutaciones confieren resistencia a todas las carboxamidas. Por ejemplo, los aislados resistentes a boscalid que presentan la mutación tirosina (H272Y) son hipersensibles a fluopiram e isofetamida. Por lo tanto, si en una población se detecta una alta frecuencia de este tipo de aislados, existen alternativas para controlarlos de manera efectiva.
SITUACIÓN ACTUAL DE SENSIBILIDAD A FENILPIRROLES (FLUDIOXONIL)
Se ha demostrado que la pérdida de sensibilidad a fludioxonil se puede originar como resultado de la sobreexpresión de transportadores multidroga en la célula fungosa. Dichos transportadores, ubicados en la membrana plasmática, eliminan o excretan el fungicida de la célula, impidiendo que ejerza su acción. El resultado es como si hubiéramos aplicado una subdosis.
A estos antecedentes se adiciona lo recientemente determinado en nuestro laboratorio, en donde al analizar molecularmente una muy pequeña población de aislados de Botrytis chilenos recuperados de uva de mesa, se detectó la presencia de una mutación en una proteína que dirige la expresión de otros genes necesarios para la sobreexpresión de los transportadores multidroga (Tesis Doctoral U. de Chile, Ch. Copier).
Si bien se ha comprobado que la generación de resistencia a fludioxonil en Botrytis tiene un costo metabólico y los aislados sobreviven menos que los sensibles, en los pocos aislados resistentes chilenos no se ha observado tal comportamiento, ya que los aislados resistentes y sensibles producen similar cantidad de esclerocios y esporas.
¿Podemos señalar que en Chile existe resistencia a fludioxonil en las poblaciones de Botrytis? No, no podemos afirmar eso. Sí podemos señalar el riesgo potencial de que ello ocurra si no se realiza un uso adecuado. ¿Por qué? A buen entendedor pocas palabras: fludioxonil está disponible en distintas formulaciones, en total actualmente serían entre 7 y 8.
Ahora bien, ¿por qué se ha incluido a fludioxonil en tantas nuevas formulaciones? Muy simple, porque quieren tenerlo como ingrediente activo en sus mezclas para contrarrestar la pérdida de sensibilidad de las poblaciones a fenhexamida y a boscalid, y porque inhibe la germinación de las conidias, inhibe la elongación del tubo germinativo y en últimos estudios recién efectuados se ha observado que también tiene un efecto interesante en etapas iniciales del crecimiento micelial de Botrytis.
Proteger esta importante herramienta de control contra la generación de resistencia es una tarea de todos, utilizándola de manera racional, con mayor razón si se usa sola, no en mezcla, en floración o en precosecha. Se deben considerar medidas antirresistencia, como aplicar un producto multisitio antes y después de fludioxonil, de preferencia extractos de plantas. Por otro lado, es importante señalar que las aplicaciones de fludioxonil en postcosecha no generan riesgo alguno de resistencia para esta u otra molécula.
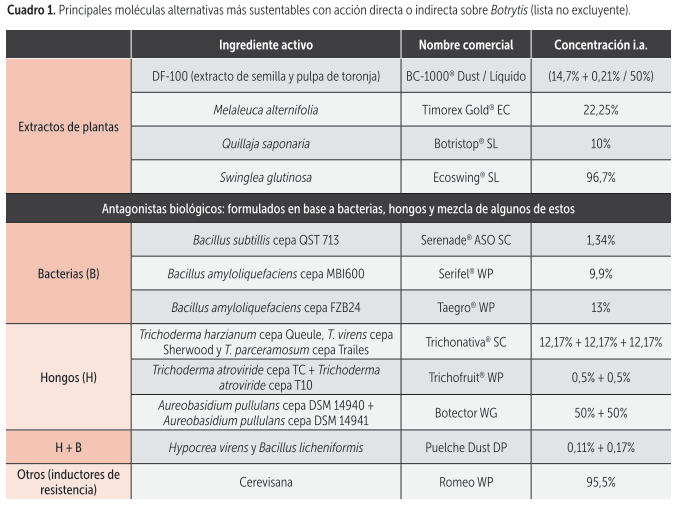
En el cuadro 1 se presentan algunas de las principales alternativas de control complementario que nos ayudan a producir de manera más sustentable (extractos de plantas, antagonistas biológicos e inductores de resistencia). Estas herramientas, aplicadas oportunamente en periodos no tan críticos, en forma previa o posterior o junto a fungicidas de síntesis, además de otorgar un control interesante, permiten llegar a destino con menor carga residual, evitar el desarrollo de nuevas resistencias y recuperar la sensibilidad a las moléculas fungicidas de síntesis que son actualmente la base de los programas de control de Botrytis.
LO FUNDAMENTAL PARA LOGRAR UVA SIN BOTRYTIS
Después de una temporada exitosa, diseñemos programas óptimos que protejan la calidad de la UVA CHILENA.
En la última temporada empezamos preocupados por una floración de muchas condiciones favorables para Botrytis y la terminamos, aparentemente, desde el punto de vista económico bastante bien en comparación a años anteriores. Lo ideal es que en la próxima temporada produzcamos “UVA PREMIUM”, con menor carga residual, pero sana, sin Botrytis, y que así la uva chilena siga siendo sinónimo de CALIDAD.
Para lograrlo, aparte de las políticas de Estado, de estrategias de marketing u otras medidas, tenemos que creernos esto. Con ese fin es fundamental diseñar programas de control adecuados a la condición de Botrytis existente en los predios, considerando lo siguiente:
1° Debemos saber qué se debe aplicar realmente, de acuerdo con el potencial de inóculo del predio y según cómo esté compuesta la población de Botrytis en cada variedad y cuartel. ¿Predominan poblaciones sensibles o resistentes a algunas de las moléculas fungicidas claves? ¿Qué tipo de mutantes predominan en la población y cuál es su frecuencia?
2° Debemos posicionar los fungicidas según condición de sensibilidad, teniendo en cuenta su efecto sobre el patógeno: si afectan la germinación de las conidias, la elongación del tubo germinativo o el crecimiento micelial.
3° Proteger los periodos críticos de infección, en floración y desde envero a precosecha es más que fundamental.
La floración requiere ser protegida en todas las variedades desde inicio a término en la zona Norte, zona Centro Norte y la zona Centro Sur. Si aplicamos en FLORACIÓN lo justo, lo más efectivo para la condición de Botrytis, el potencial de infección en envero y precosecha será más bajo y por lo tanto más fácil de controlar. En Cambio, si el programa de control de flor no fue el adecuado… programas baratos en flor al final cuestan muy caro.
