



Interacción planta-microorganismo
Importancia de los exudados de las raíces en el diálogo entre Trichoderma y las plantas
Las plantas, como productoras primarias de ecosistemas, albergan una infinidad de especies microbianas, que dependen de sustancias ricas en carbono exudadas por las raíces para su nutrición, como azúcares, ácidos orgánicos y aminoácidos. Gran parte del microbioma fúngico y bacteriano permanece neutral, pero unas pocas especies pueden establecer relaciones patógenas o simbióticas que, en última instancia, influyen en la capacidad de adaptación y productividad de las plantas (1). Uno de los principales objetivos de la gestión sostenible de cultivos es identificar y caracterizar especies microbianas con rasgos probióticos. Estos últimos permitirán detectar sustancias altamente activas con el fin de desarrollar una nueva generación de pesticidas, bioestimulantes y elicitores de defensa para un mercado en crecimiento. En este escenario, el género fúngico Trichoderma está alcanzando una importancia cada vez mayor ya que consta de más de 200 especies reportadas con metabolismo versátil y propiedades bien adaptadas para proliferar en ambientes de suelo y agua (2).
24 de Junio 2021
Equipo Redagrícola

Sarai Esparza-Reynoso y José López-Bucio, Instituto de Investigaciones Químico-Biológicas, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México.
email: sariesparza@gmail.com ; jbucio@umich.mxt
Gracias a las investigaciones realizadas en los últimos veinte años, hemos podido conocer mejor los efectos beneficiosos de Trichoderma en las plantas respecto a: 1) ramificación de las raíces y potencial de absorción, 2) uso de enmiendas orgánicas y fertilizantes, 3) crecimiento y desarrollo, y 4) adaptación a los desafíos abióticos y bióticos. En resumen, muchas especies, como T. virens, T. atroviride y T. longibrachiatum, pueden ayudar a las plantas a sobrevivir mejor y a aumentar la productividad de forma segura y ecológica. Se ha evaluado el impacto de estos hongos en condiciones de campo al obtenerse un aumento del rendimiento en importantes cultivos de cereales, frutas y verduras, como maíz, trigo, soja, tomate, uva y lechuga (2). Una de las principales interrogantes de los últimos años ha sido la forma en que Trichoderma adapta su metabolismo en función de los nichos ecológicos y los recursos nutricionales altamente variables que encuentra. Aparentemente, la secreción de enzimas potentes, como celulasas, quitinasas y peptidasas, son el sello distintivo que permite la explotación de madera muerta y materiales de hojas, raíces y tallos en descomposición y subraya su capacidad para parasitar hongos fitopatógenos. Sin embargo, al parecer la represión de los genes que codifican las enzimas degradativas de los hongos permite la colonización de las raíces por parte de Trichoderma y, por tanto, el hongo reconoce las raíces sanas por medio de sus patrones de exudación (3).
DECODIFICACIÓN DEL MENSAJE QUÍMICO DE TRICHODERMA
Trichoderma [hongos] viene a ser una fábrica de sustancias orgánicas que libera compuestos volátiles, hormonas vegetales, metabolitos secundarios y pequeños péptidos cuya composición molecular depende de varios factores, como la especie de hongo, la disponibilidad de nutrientes y la interacción con microorganismos y plantas. Estos infoquímicos pueden ser percibidos por las raíces mediante la difusión libre dentro del suelo y la materia orgánica, así como a través del contacto físico entre las hifas y la epidermis de la raíz, o en etapas posteriores, donde el hongo se extiende a las células corticales internas (4-6). El primer cambio manifiesto en la rizósfera como consecuencia de la presencia de Trichoderma es la acidificación del pH (7). Esto puede explicar su alto rendimiento en la solubilización de fosfatos poco solubles, lo que representa una mejor nutrición de la planta. A medida que el hongo crece, se cree que los compuestos volátiles sensibilizan a las raíces y permiten el reconocimiento a larga distancia entre raíz y hongo. La 6-pentyl-2H-pyran-2-ona (6-PP) es el principal compuesto volátil de las mezclas de T. atroviride que desencadena la ramificación de las raíces en arabidopsis a través de cambios en el transporte y la respuesta de la auxina y el etileno, respectivamente (4). Se ha descubierto que el alqueno 1-deceno reprime los genes de defensa, de estrés y de respuesta a las enfermedades, lo que facilita la propagación de hongos en los tejidos de las raíces (6). T. virens y T. asperellum liberan auxinas, un tipo de fitohormonas con funciones en el crecimiento e inmunidad de las plantas, que pueden estar directamente relacionadas con sus propiedades bioestimulantes (8, 9). El reconocimiento físico puede desencadenar otras reacciones tanto en el hongo como en la planta. La quitina, uno de los principales componentes de las paredes celulares de los hongos, se considera desde hace tiempo un elicitor que desencadena reacciones defensivas en las plantas. Existen otras moléculas, tales como pequeños péptidos y proteínas fúngicas de la membrana o del citoplasma, que pueden alertar a las raíces a prepararse para la interacción con el fin de evitar efectos nocivos, haciéndolas mucho más competitivas (10). A través de la proliferación de raíces laterales y adventicias, las plantas aprovechan mejor los recursos minerales e hídricos y son más resistentes al estrés abiótico, procesos que son inducidos eficazmente por Trichoderma. Las compensaciones entre crecimiento y defensa en las plantas se regulan durante las interacciones bióticas. Esto permite que las transiciones de desarrollo hacia la floración se lleven a cabo y garanticen la producción de semillas. Dado que Trichoderma desencadena tanto la inmunidad de la planta como su crecimiento, es difícil determinar si estos mecanismos, que son intrínsecamente difíciles, puedan estar relacionados. Más aún, cabe la posibilidad de que las raíces colonizadas por el hongo tengan un mayor metabolismo para soportar la demanda energética de la simbiosis. Se ha descubierto que la exudación de carbohidratos a través de las raíces de plántulas de tomate incrementa su respuesta a T. atroviride y que los azúcares pueden actuar como una respuesta quimiotáctica positiva para atraer a las hifas (11), como se indica en la Figura 1. En consecuencia, el hongo expresa una invertasa intracelular (Triat51014) y dos transportadores putativos de sacarosa (Triat226844 y Triat83012) para utilizar el carbono derivado de las plantas. Por otro lado, T. virens capta la sacarosa a través de un transportador de sacarosa y la hidroliza a través de una invertasa intracelular, lo que permite un rápido crecimiento gracias a la energía proporcionada por este disacárido (12).
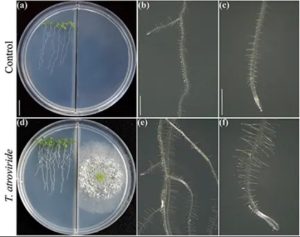
Las pruebas moleculares ayudan a explicar cómo Trichoderma cambia sus firmas metabólicas al detectar exudados radiculares. Villalobos-Escobedo y cols. (2020) fueron más allá y demostraron que la expresión de los genes que codifican las enzimas implicadas en la degradación de carbohidratos complejos, como la celulosa o la quitina, antes de la colonización de las raíces está comprometida en los mutantes de T. atroviride defectuosos en NADPH oxidasa, una enzima directamente implicada en la producción de especies reactivas de oxígeno. La mutación de los genes correspondientes no sólo afecta la formación de raíces laterales y la producción de biomasa en la planta huésped, sino que también afecta las reacciones de defensa provocadas y el comportamiento saprófito del hongo para adquirir y utilizar los azúcares simples disponibles en los exudados de las raíces (3). Se ha discutido mucho si acaso los rasgos de las plantas que subyacen a la alta producción de biomasa en las interacciones con los microbios dependen esencialmente del dióxido de carbono emitido. Ciertamente, el dióxido de carbono es uno de los principales reactivos de la fotosíntesis y debiera explicar el aumento de la biomasa. Sin embargo, la estrecha comunicación depende de la composición molecular de las mezclas volátiles. Al probarse el crecimiento y desarrollo de la planta en placas Petri divididas para evaluar la interacción hongo-planta a través de los volátiles (Fig. 2), hemos revelado el papel crítico de la 6-PP en la producción de biomasa de la planta que influye directamente en las respuestas hormonales, y el transporte de azúcares a larga distancia entre el brote y la raíz (Esparza-Reynoso y cols. presentado). Esto respalda la noción de que el estado del carbono en el brote y la percepción de los metabolitos fúngicos son críticos para la mitosis en los meristemas de las raíces cuando un bucle de regulación para la distribución de azúcares depende de la fotosíntesis. Curiosamente, los hongos promotores del crecimiento de plantas, como las cepas de Trichoderma y Serendipita indica y S. williamsii, promueven fuertemente la fotosíntesis a través de mezclas de compuestos volátiles (13, 14) y no de compuestos individuales. La inhibición por retroalimentación de la fijación de dióxido de carbono provocada por los elevados niveles de azúcar aparentemente no se produce en las hojas expuestas a los volátiles fúngicos, posiblemente debido a un desequilibrio hormonal. Las plantas no están solas, dependen de sus simbiontes fúngicos para sobrevivir y prosperar. Ha llegado el momento de que los conocimientos actuales se materialicen en la aplicación de Trichoderma en el campo.
Las compensaciones entre crecimiento y defensa en las plantas se regulan durante las interacciones bióticas.
Esto permite que las transiciones de desarrollo hacia la floración se lleven a cabo y garanticen la producción de semillas. Dado que Trichoderma desencadena tanto la inmunidad de la planta como su crecimiento, es difícil determinar si estos mecanismos, que son intrínsecamente difíciles, puedan estar relacionados.
Referencias
- Cordovez V, Dini-Andreote F, Carrión VJ, Raaijmakers JM (2019). Ecology and evolution of plant microbiomes. Rev. Microbiol. 73:69-88.
- López-Bucio J, Pelagio-Flores R, Herrera-Estrella A (2015). Trichoderma as biostimulant: exploiting the multilevel properties of a plant beneficial fungus. Hort. 196:109-123.
- Villalobos-Escobedo JM, Esparza-Reynoso S, Pelagio-Flores R, López-Ramírez F, Ruiz-Herrera LF, López-Bucio J, Herrera-Estrella A (2020). The fungal NADPH oxidase is an essential element for the molecular dialog between Trichoderma and Arabidopsis. Plant J. 103:2178-2192.
- Garnica-Vergara A, Barrera-Ortiz S, Muñoz-Parra E, Raya-González J, Méndez-Bravo A, Macías-Rodríguez L, Ruiz-Herrera LF, López-Bucio J (2016). The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning. New Phytol. 209:1496-1512.
- Lee S, Behringer G, Hung R, Bennett J (2019). Effects of fungal volatile organic compounds on Arabidopsis thaliana growth and gene expression. Fungal Ecol. 37:1-9.
- Guo Y, Ghirardo A, Weber B, Schnitzler JP, Benz JP, Rosenkranz M (2019). Trichoderma species differ in their volatile profiles and in antagonism toward ectomycorrhiza Laccaria bicolor. Microbiol. 10:891.
- Pelagio-Flores R, Esparza-Reynoso S, Garnica-Vergara A, López-Bucio J, Herrera-Estrella A (2017). Trichoderma-induced acidification is an early trigger for changes in Arabidopsis root growth and determines fungal phytostimulation. Plant Sci. 8:822.
- Contreras-Cornejo HA, Macías-Rodríguez L, Cortés-Penagos C, López-Bucio J (2009). Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 149:1579-1592.
- Wang YF, Hou XY, Deng JJ, Yao ZH, Lyu MM, Zhang RS (2020). AUXIN RESPONSE FACTOR 1 acts as a positive regulator in the response of Poplar to Trichoderma asperellum inoculation in overexpressing plants. Plants (Basel) 9:272.
- Rocafort M, Fudal I, Mesarich CH (2020). Apoplastic effector proteins of plant-associated fungi and oomycetes. Opin. Plant Biol. 56:9-19.
- Macías-Rodríguez L, Guzmán-Gómez A, García-Juárez P, Contreras-Cornejo HA (2018). Trichoderma atroviride promotes tomato development and alters the root exudation of carbohydrates, which stimulates fungal growth and the biocontrol of the phytopathogen Phytophthora cinnamomi in a tripartite interaction system. FEMS Microbiol. Ecol. 94:fiy137.
- Vargas WA, Mandawe JC, Kenerley CM (2009). Plant-derived sucrose is a key element in the symbiotic association between Trichoderma virens and maize plants. Plant Physiol. 151:792-808.
- Harman GE, Doni F, Khadka RB, Uphoff N (2019). Endophytic strains of Trichoderma increase plants’ photosynthetic capability. Appl. Microbiol. https://doi.org/10.1111/jam.14368.
- Venneman J. et al. (2020). Respiratory CO2 combined with a blend of volatiles emitted by endophytic Serendipita strains strongly stimulate growth of Arabidopsis implicating auxin and cytokinin signaling. Plant Sci. https://doi.org/10.3389/fpls.2020.544435.
